
How Did Such A Risky Thing As The Imagination Evolve?
The engine of Social Space is the human imagination. It is the evolution of this ‘engine’, along with the attendant development of language, that marks the change from Animal to Man. Other candidates for this change from Nature to Culture have been considered in the past, such as our bipedal stance, opposable thumbs and extended infancy. But these factors could not, at least on their own, lift man above the strict causal framework of animal existence. Nor was it the use of fire, the creation of tools, nor even the invention of soft toilet rolls that gave our race its proven ability to soar above the strictures of Physical and Genetic Space. Because all these factors are just pieces that play their part in a much bigger picture, or rather a much greater storyboard. A storyboard, that centres around the evolution of the imagination and the development of language. For these are the two factors responsible for the emergence of the human species as a radically new form of life on earth. And of the two, it is the evolution of the imagination that will mainly concern us here.
What then is so special about the imagination? What new ability does it bring to the arena of animal nature? What is the basis of its power? Well, probably the best way of looking at this is to go back to our roots in the evolution of animal behaviour. There, we can we find out whether there really is an important difference between the rest of the animal kingdom and ourselves, and identify what that difference might be.
So, one interesting way to look at the nature of this new power might be in terms of how both the animal kingdom and the human race are synced into the past, the present and the future. What we would see in that case is that whilst the animal kingdom is firmly fixed to the present and tightly tied to the past, the human race is free to consider the future, and reflect on what might be. Or to put this another way, if animal behaviour was looked at as a form of grammar, it would surely divide into these three tenses:
The Sensory Present (how things are right now, ‘before our very eyes’)
The Individual Past (the ‘ontogenetic perfect’ of how things have transpired in the past of each individual)
The Species Past (the ‘phylogenetic perfect’ of how things have transpired in the past of the species).
The question is, where would we put the imagination in the context of such a ‘grammar’, and in particular, where does it belong alongside these three ‘tenses’ of animal behaviour? Well, the answer seems to be that the human imagination adds a whole new tense to this ‘animal grammar’, namely, the future or subjunctive tense.
The Imagined Future (the subjunctive tense of ‘what might be the case’ in the future).
So how did this new and potentially dangerous speculative tense evolve? The answer presumably lies in an understanding of how the other three ‘tenses’ evolved in the first place. That is, after all, where the imagination first emerged from, to then consolidate its position as a new and revolutionary fix on reality, so that is where we must go, to understand its evolutionary history. And to do this, we can take a quick look at what we might call the more conventional ‘orthodox animal grammar’ by way of a few simple examples.
When a fledgling bird crouches low at the sight of a hawk flying overhead, it is using two tenses in its response to this predator – ‘the sensory present’ and ‘the species past’. So when the fledgling sees the hawk, its eyes transfer a real-time picture of ‘the sensory present’ to its central nervous system, where it is then matched with a picture from ‘the species past’. That is, the chick is programmed to scan the present, and compare the results against the programming of the ‘species memory’. Almost as if the species memory carries an innate ‘picture’ of an aerial predator silhouette (held there as the result of past selection pressures), which the fledgling can match against the incoming picture from its eyes. And if the two match, an inbuilt response is triggered, and the fledgling crouches.
In other words, we are looking at an innate pattern of both recognition and response from the species past, which is then matched against an active real-time picture from the sensory present. We can call this ‘picture matching’ (and this just happens to be a metaphor that serves our purpose here, as the mechanism is not the issue in this comparison). So, when the species past and the sensory present are used as two streams of data that are then compared with each other, then for the purpose of this section, that is what is meant by the term ‘picture matching’
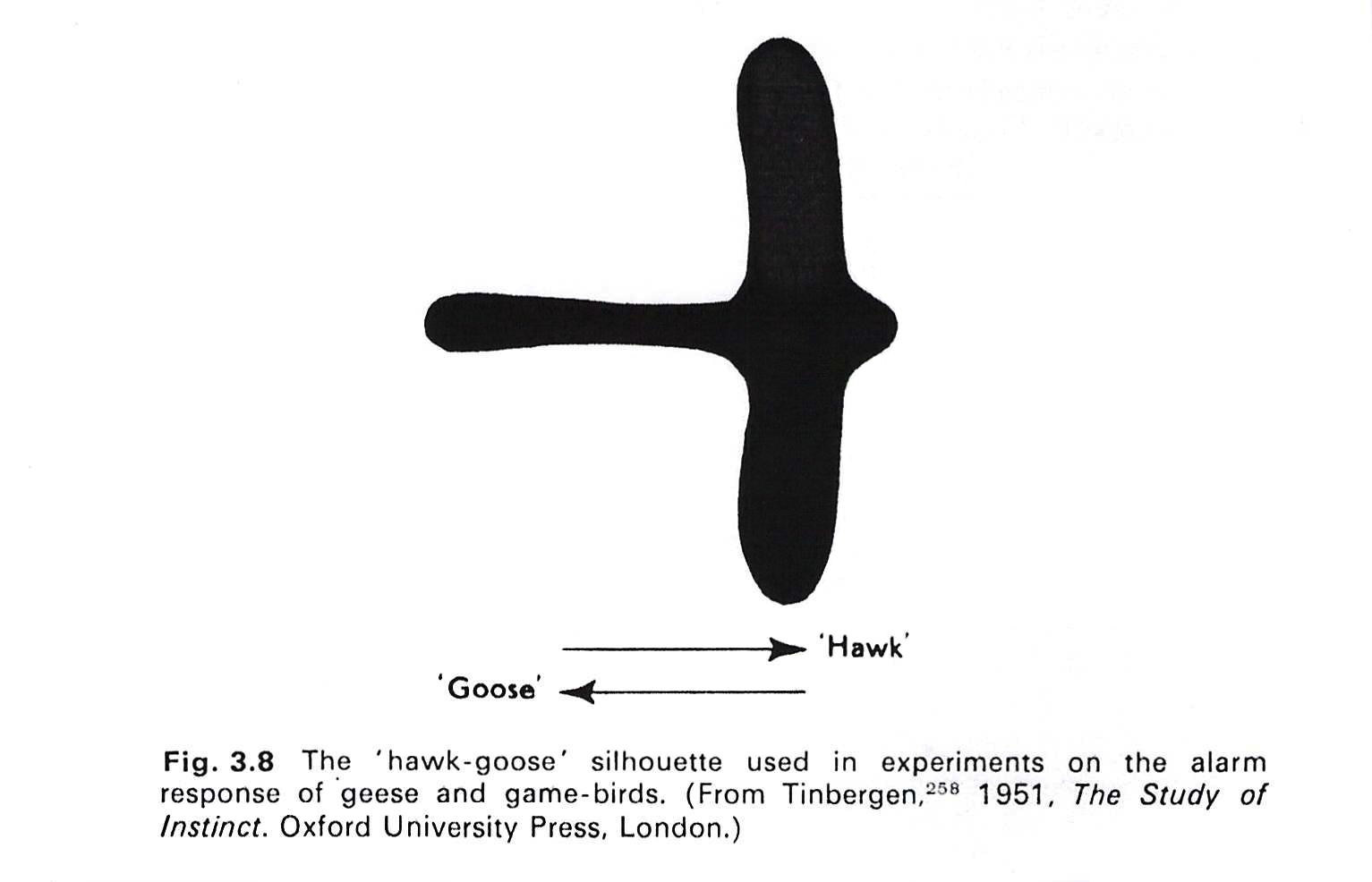
Below is a visual construct used in a well known experiment that explored this particular crouching response of fledgling birds on the ground.
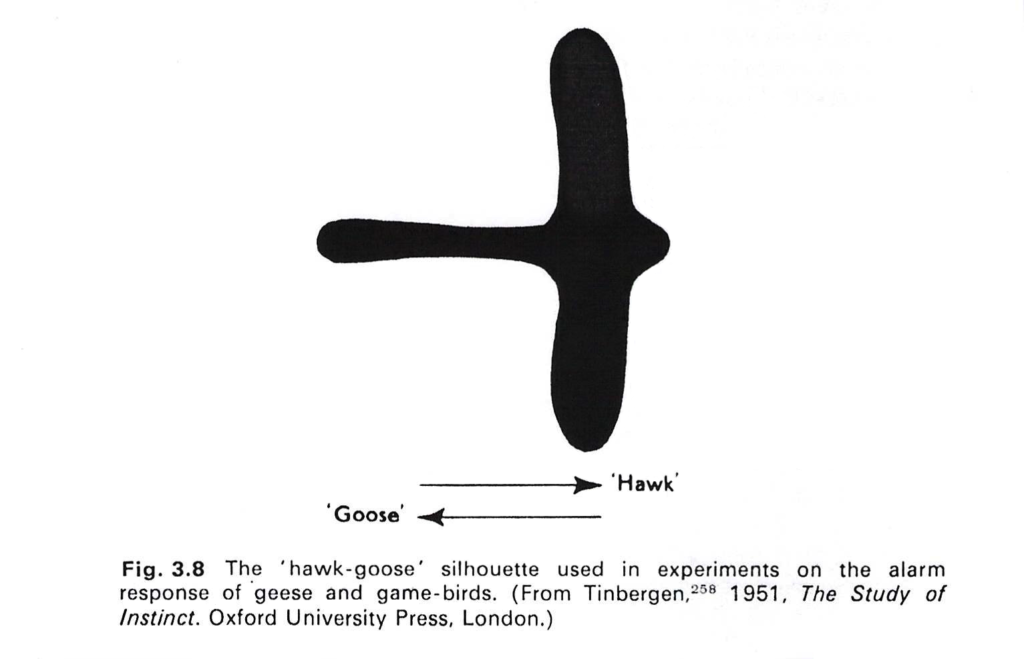
Interestingly, it seems from the experiments performed by Tinbergen and others, that a silhouette pulled across the chicks in one direction elicits the crouching response, but the chicks ignore it when it is pulled across in the opposite direction, where it looks more like a goose than a hawk. It also seems that the slow flapping flight of the hawk or buzzard is more likely to trigger the behaviour than a faster flight pattern. Either way, the point is that the fledgling is responding to its species past when it sees a hawk fly above it, by matching its species memory to its own real time sensory picture in the present.
In fact, this system of instinctive response is so effective that it is hard to see why other tenses of animal grammar then evolved. Surely, one could argue, the prior knowledge that is entailed in the innate recognitions and responses of the species past must always be superior to anything else? Because this way, the fledgling bird can avoid learning about hawks the hard way, thus avoiding any real and dangerous exposure to the so called ‘school of hard knocks’, which in this particular case could mean getting eaten. So the bird survives thanks to to the programming evolved during the past of its species. Which is to say that as long as the sensory present can match pictures against the key pictures picked up by its phylogenetic past, the successful responses can be triggered, and the animal will live to crouch another day. After all, this is the only thing that counts in Genetic Space; because then the response will be carried onto the next generation, and its holder will survive to promulgate its genes further. All of which leads us to our next question. If being wise before the event is such an evolutionary winner, why on earth did learning, with its new and relatively short-lived tense of the individual past, evolve at all? Or to put it another way, how on earth can being wise after the event help an animal to increase its chances of survival?
‘Being wise after the event’ is an expression that is often preceded by the words: ‘Well, it’s all very well being…’ Meaning that it is better to be wise before things happen rather than after them, especially if ignorance of the event leads to damage or death. But in other circumstances, where delay or ignorance is not immediately lethal, the phrase ‘better late than never’ may apply. Meaning that learning (as opposed to knowing) about something can be useful next time round – as long as there is a next time round…
As an example of learned behaviour, let us look at the Digger Wasp, and its ability to learn and recognise the whereabouts of its nest site. The problem in this case being that although a digger wasp must have an innate directive to seek out a sandbank for its burrow, there is no way that its genes can provide the specific details of an actual site because there is no way of knowing such details in advance. Only by actively learning such details can the wasp then locate its burrow site when it returns with the food for its young. And it is in conditions like these that the capacity to learn must gain a significant selective advantage in the repertoire of an animal. Because if the wasp can quickly update on the shifting sands of its own environment, and within the brief span of its adult life, rather than relying on the much slower timescale of genetic evolution, then it can overcome the limitations written into the innate species response, and survive. Which is why learning is such a common strategy in the animal kingdom. Because in matters that are unpredictable, such as the look of a place, or the change in behaviour of a predator, then having an individual memory is the only solution to the shortcomings of species memory.
Below we see one of the diagrams illustrating an experiment that was designed to confuse the Digger Wasp, by moving the goal posts as it were. In this case by moving the pine cones to show how the wasp relies on the cones as landmarks for the relocation of its burrow.

In this experiment, it is clear that the digger wasp checks out the landmarks around its nest in real time, before then moving off to find a caterpillar and bring it back. It also demonstrates how even learnt behaviour cannot cope with sudden changes, such as those brought about by the arrival of a band of investigative scientists. True, the wasp will eventually find its nest burrow again, and update its navigational images accordingly, so it is not the end of the story, but it does suggest that learning shows the same kind of limitation as the longer term species programmed response pattern. Namely, that both can be forced out of date by a changing environment.
As with the crouching chick example, it is interesting to look at the digger wasp example in terms of ‘picture matching’. We have seen that in the case of the chicks, their instinctive response relies on picture matching between the sensory present and the species past, so what does the mechanism of learning involve? The answer lies in a modification to this already successful system. A modification that works by way of a ‘selective freezing’ of the real time pictures coming in from the sensory present. The point being that if these pictures can be selectively ‘frozen’, then this new information can be stored and matched against further incoming sensory data in just the same way that matching occurs in instinctive behaviour. For example, once the picture of the territory surrounding the digger wasp site has been frozen in memory, its identity can be matched with further pictures coming in from the eyes of the wasp as it comes home. And the match between the frozen and real-time pictures will enable it to check and correct its course back to the burrow. So what we see here is that the phylogenetically old and reliable system of picture matching has been tweaked to achieve a dynamic new set of possibilities. The result of which is a major development in behavioural evolution. For now the animal is no longer bound by the past of its species – it can learn new things for itself, and adapt to its changing environment in the real time of its own individual present.
All of which leads us to pose the following question. If it is true that the dual strategy of Innate Knowledge and Individual Learning add up to such a winning combination, then what possible advantage is there to be gained by evolving the further, and dangerously speculative tense of the so called ‘subjunctive mood’ of ‘what might be’?
Again, the answer comes down to a question of limitations. The thing is, learning does bring with it certain major advantages, but in some ways it too can fall short under certain circumstances. For example, if an animal faces a brand new and dangerous situation, where the cost of getting it wrong is death, then the price of learning this lesson from scratch is certainly to be avoided. But how can the animal avoid this dilemma? Well, in principle, an innate response from the species memory could get around this problem, except that in this case, we have described the situation as ‘brand new’. Which means there has been no time for the new behaviour to arise by chance mutation, let alone get fixed in the gene pool. So neither learnt nor evolved behaviour patterns are adequate in this situation, because neither are up to this confrontation with the brand new circumstances that can suddenly occur in the lifetime of an individual. So the question is this: How can an animal learn new things, during its own lifetime, whilst avoiding the potential cost of that learning?
One answer is that the animal can remove itself from the direct consequences of error by spotting the mistakes made by others, or by imitating the behaviour of fellow members of its group that have somehow learnt to avoid it. That is, in principle, it could learn from its parents or other conspecifics by copying. An opportunist strategy, as it were, that places the user one step away from the line of fire. Vicarious perhaps, but effective. Could this then be the answer to our problem of the dangers of trial and error? Perhaps so. After all, the strategy of imitation does work in certain situations, and it does sidestep certain problems. Well, just as long as the other members of the group are able to meet the challenges around them that is. But that is not always the case. Or, they may not easily share the advantage they have acquired. Or perhaps they are unable to pass it on because they are not there when the danger presents itself. Or the species is solitary, like the tiger. So the fundamental question remains. Is there any way in which the challenge of the environment can be answered without the limitations and dangers of imitation and learning?
Consider then the behaviour of a chimp in an experimental situation where the chimp is separated from a test (along with the inevitable banana) by a set of bars. The bars ensure that it sits on the wrong side of the coveted banana, giving it observer status only. And from its vantage point, it sees there is only one way to reach this banana. Namely, through the clever combination of objects that have been left on the floor inside the cage where the banana happens to be. So, to reach its prize, the chimp must reach through the grill for a stick that is, in itself, too short to reach the banana. But what it can do is use it to reach another longer stick, and then use that one to gain its reward. Now, not surprisingly, chimps generally accomplish this task with ease. But what sort of mental process are they using to solve this problem? Is it just another case of learnt behaviour, or is a different order of mental process involved?

In the accounts of such experiments, it is apparently common for the chimp to pause, and stare around somewhat vacantly, between an initial failure to secure the banana, and a sudden resolution of the problem. An interval of hesitation and doubt that possibly marks what could be an internal attempt to float various possible combinations of the objects on the other side of the grill, always with the banana firmly in mind. The chimp gazes round, juggles these different alternatives in its head, arrives at a solution, acts upon it, and gets the banana. Now, it seems likely that such a juggling requires an internal modelling of reality qualitatively different to the animal grammar we have considered so far. Because this is a new form of mechanism that leads to an alternative view of reality that the chimp has never come across before – in this case concerning the two sticks. The point is that such a new combination may well represent a radical departure from the high-fidelity picture-making and picture matching that is characteristic of normal animal behaviour. So what is the nature of this departure, and what gives this new behaviour such a selective advantage?
Innate and learnt behaviours are inherently conservative because they simply can’t afford to play around with any biological facts that may mean the difference between life and death. Which is why animal behaviour is doggedly faithful to both the ‘what has been’ of the species past, the ‘what has more recently been’ of the individual past, and the ‘what is going on right now’ of the real time sensory present. And why is this fidelity so critical? Well, one has only to look at what happens when innate, learnt and real-time pictures are disturbed by forces such as mutation, memory lapse, or poor sensory reception to find the answer. Basically, an animal that stops being faithful to its present and past is not going to survive for long.
However, the imagination is not locked to the past in this way. In fact, in contrast to these inborn and learnt behaviours, it is inherently creative. So it unfreezes the existing picture-making ability of the brain, freeing it from its literal-minded fidelity to the past and present. In so doing, it floats these elements to the surface of an altogether new, and powerful level of awareness. A level where these pictures can be held and realigned with the aim of creating new combinations and new possibilities. But we do have to ‘keep it real’ somehow. Because if we leave reality too far behind, it is going to catch up with us, and pull us down to earth with a lethal bump. So our new set of possibilities must always remain consistent with the parameters set up by the reality outside us (at least in its early stages of evolution), or our ‘new awareness’ will lead us into danger, potential damage and even death.
To help ‘keep things real’, the new tense of ‘What might be’ must be closely channelled by the instinctual drives of the animal. These drives are what gives the imagination its goals after all. Meanwhile, the senses must continue to present the brain with an accurate picture of what is going on outside, and the memory must continue to supply the individual with its past experience, because it is between the real time present and the individual past that the imagination is going to float a choice of likely combinations and solutions. For example, our chimp, when faced with a problem that cannot be solved through innate or learnt abilities, momentarily frees its mind from this slavish alliance to the world of ‘what has been’ and the fixity of ‘what is’, and instead pictures the world of ‘what might be’. And it is only by doing this that the chimp is able to circumvent the limitations of standard animal response, and create the radically new set of responses that then sets its species a distance apart from the rest of the animal kingdom. So how then does the chimp float these new pictures from this dual input? This is a major and fundamental question facing the neurophysiologist, and the answer will surely be as fascinating as it is complicated. But the chief concern here is not how the imagination works (‘picture matching’ and ‘the subjunctive tense’ are merely metaphors after all), but rather to emphasize how the evolution of this power is fundamental to the development of the human race.
So far, we have discussed the freedoms and limitations of pre programmed and learnt behaviours. Clearly, both have major advantages, such as the instantly available knowledge that is supplied by ones ancestors, and the ability to add brand new knowledge that comes from an individual life experience. And clearly both have limitations, such as inflexibility and the dangers that come from trial and error. So what of the imagination? What advantage does it confer on its user, and what if any are its limitations?
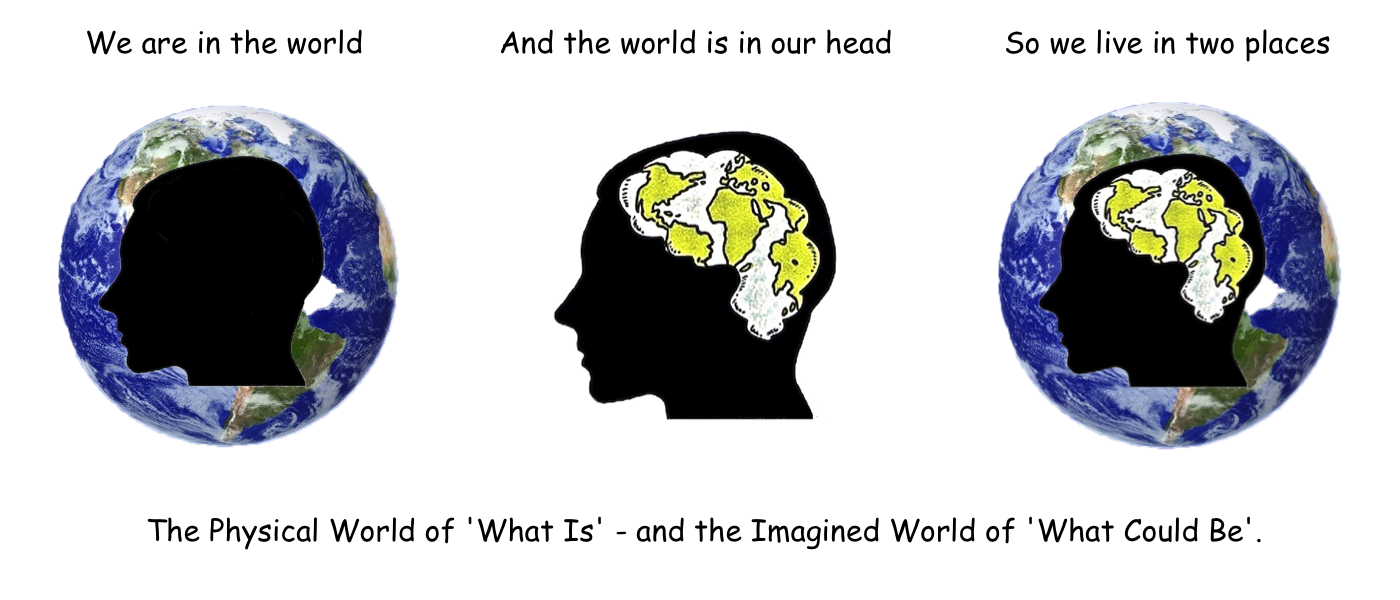
We have seen that learnt behaviour is quite flexible when compared to innate behaviour, but it still depends on direct experience for its progress in solving problems. You have to be there ‘in the Danger Zone’ to pick up the necessary information that learning depends upon. But this limitation does not apply to the use of the imagination, which can operate inside the head of its owner, and at one step removed from physical reality. A single step that makes all the difference precisely because it takes us out of the real world, and puts us into another much safer one – a virtual arena where anything goes, and where physical reality no longer rules. And it is in this new virtual world that new combinations of actions can be tried out without any risk, and new alternatives can be played out in all their possible combinations, free of both time and space. Almost as if the whole world could be remodelled inside the skull. Which is exactly the point of the first cartoon on the first page of the introduction to this website. A profile of a human head, showing our planet inside its confines, complete with all its continents, and shaped in the convoluted form of a brain. ‘We are in the world, and the world is in our head’. In fact, given its clear application to the world of what is, and what could be, here it is again.
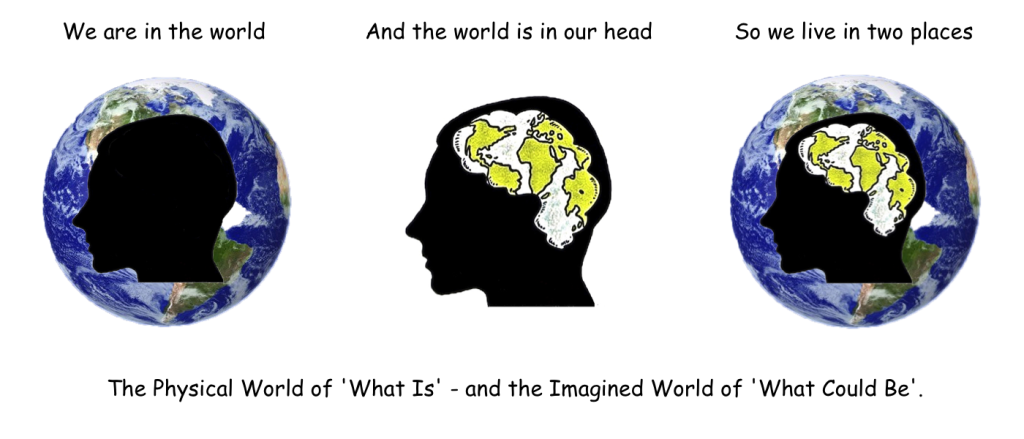
What we can say here is that the subjunctive tense of ‘what might be’ has led to that other kind of ‘might’: the Might of the human race in its dominion over this planet (it looks as if both usages come from the same Germanic origin incidentally). Because, more than anything else, it is our ability to imagine what might be that has given us the great powers that we now hold over the natural world around us. A ‘Might’ almost entirely based on our sophisticated use of the virtual reality inside our heads. However, it may not be enough to have an imagination on its own, because unless its holder has a way to communicate with the other imaginations around it, its ideas are likely to only live as long as the brain of its maker.
In particular, the human imagination needs two special agents in order to work effectively on the part of its owner. The first of these is a physical agent that will convert its virtual output into the physical action so important to achieve success in the real world, and that physical agent is the hand. The second is also a physical agent, but it converts its virtual output into the language so important to achieve success in the social world, and that physical agent is the mouth. Now clearly the hand and the mouth are only part of the whole story (they both require special areas in the brain, as well as other parts of the body for physical action and language to work), but they make perfect icons for the two conditions necessary for the imagination to then develop and increase its very real might.
The evolution of a pair of hands, with their opposable thumbs for the manipulation of the physical world, has to be a primary evolutionary requirement if the imagination is to be an effective force in nature. This is well recognised in the idea that however clever dolphins might be, they could never have dominated the planet because they have no way of manipulating the physical world. A pair of hands, it seems, is a vital tool through which the imagination can develop, and realise its true potential in the physical world. And the same goes for the evolution of the mouth (along with the voice box, and ears, and special areas of the brain). Because by using the mouth, the imagination is able to exchange and test out its ideas with other imaginations through the use of language, and it can then pass these ideas and findings on to the next generation, so that they in turn can build on them, and go further still. Words don’t just serve as a means of transmission therefore; but also as a means of cross fertilisation whereby new ideas may emerge out of the exchange of old ideas. But before the time of words, and before language evolved and developed, there was probably not a lot to talk about. Because before language, the mind as we know it was surely so embryonic that its fleeting consciousness and its intellect really did have little to impart. Well, it’s a nice idea, and it emphasizes the importance of the word: before language, there was nothing to say.
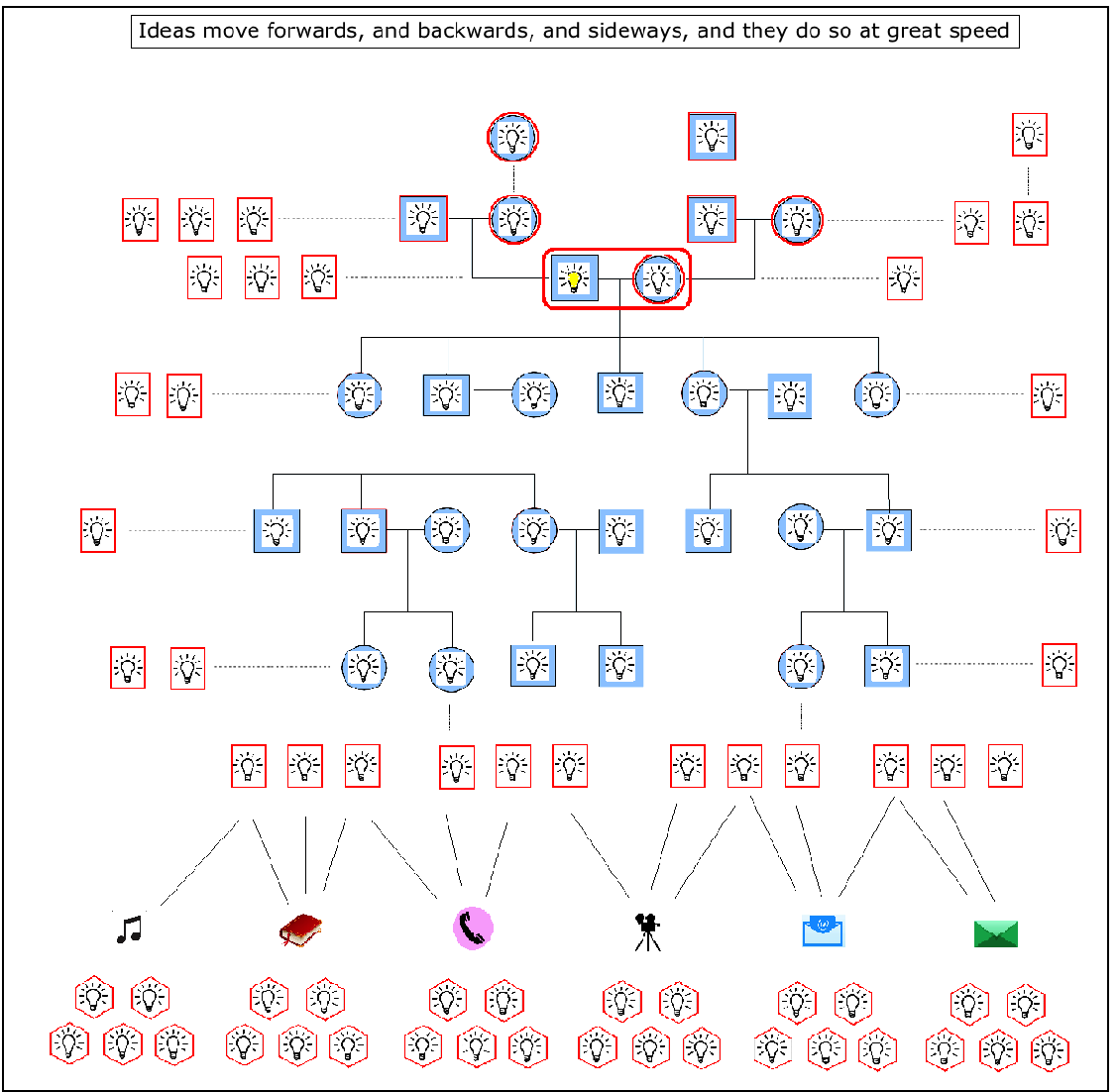
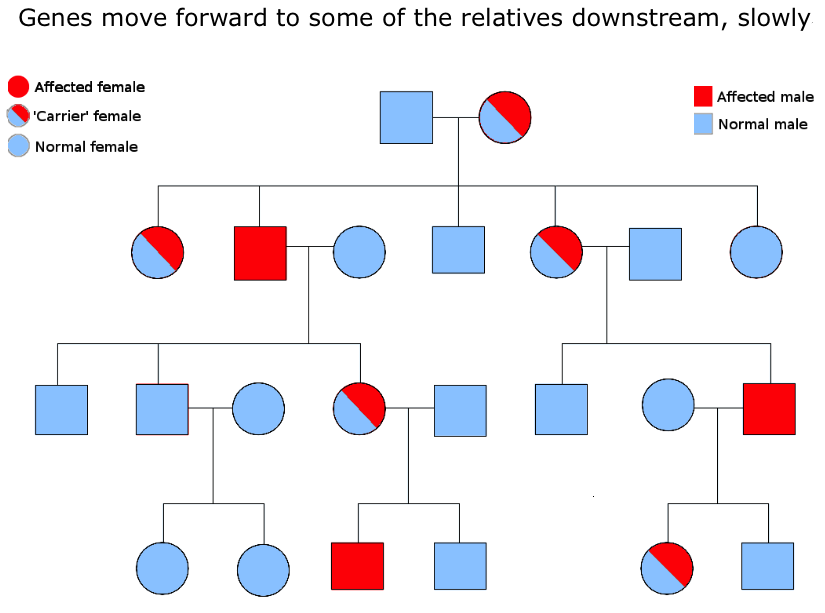
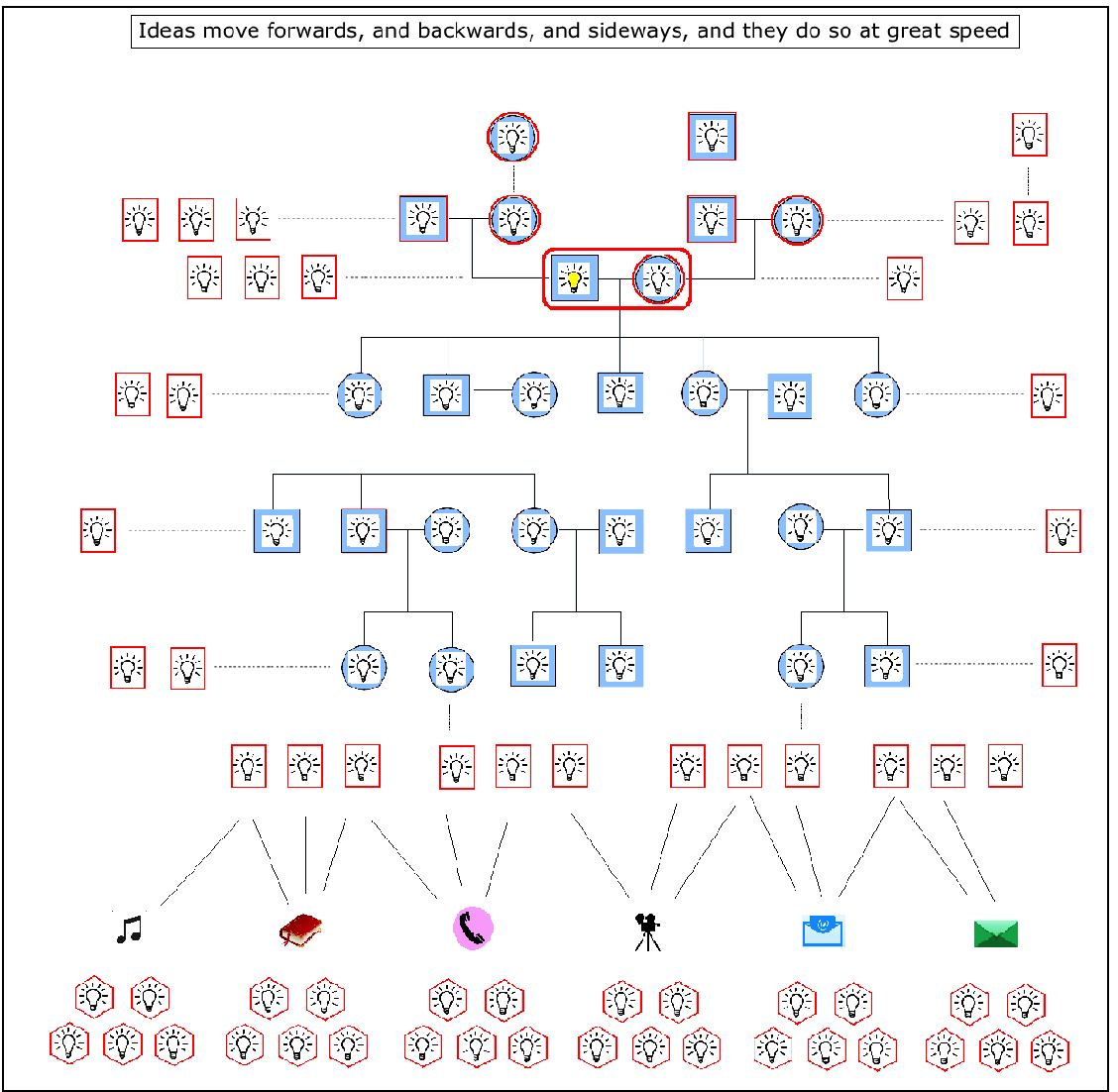
Ideas can be compared to genes in this respect. Genes have the potential to spread out downstream through successive generations, but ideas can transmit much more quickly, with wider spread, and in all directions. For example, ideas, through language, can go backwards, following upstream to the grandparents generation. They can also step sideways, to their peers in the same generation. And like genes, they can also spread out downstream, but unlike genes, which are limited to a line or increasing cone of descendents, ideas can spread out to become a veritable horizontal wave down the generations. And the reason for this major difference in freedoms? Well, ideas do not require sex amongst willing participants for their movement. So when they move downstream, through the generations, they are not limited by the major investment required by genetic transmission – because the words that carry them don’t need to bother with mate location, courtship, mating, conception and birth. They simply spread out, unhindered by genetic lines of biological inheritance, even switching languages, as if they were jumping genes moving from one species to another.
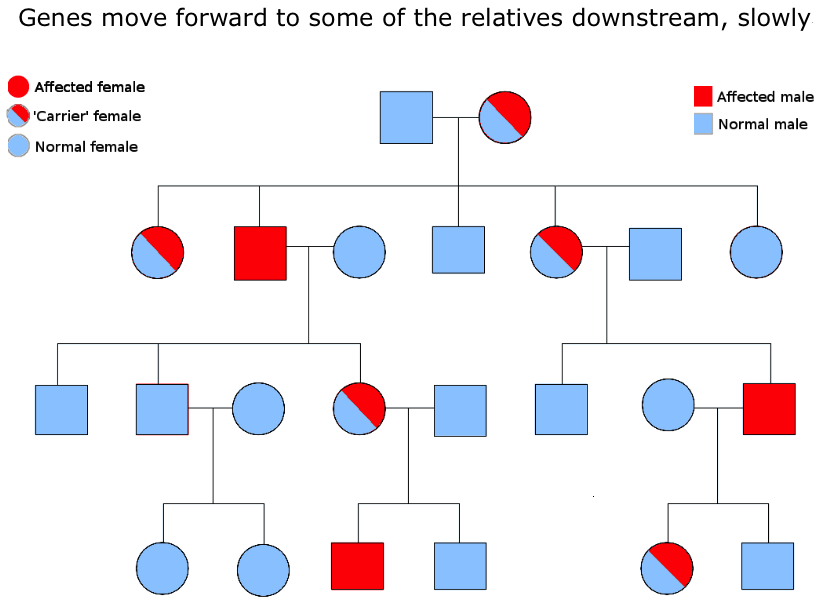
Nor does it stop there. Ideas, through language, have then gone on, in many cultures, to transcend oral culture itself. Sounds have been turned into signs and letters, and these have been transcribed into alphabets and then books, and then recorded in digital forms, such as films and pictures and designs and plans and maps, and all this has then moved into a different virtual reality from the human imagination in the form of cyberspace and the internet. Which means that human ideas can then lie dormant, or be transmitted at amazing speeds, or be used as algos and apps to make even more complex systems of thought, and finally to create new intelligence.
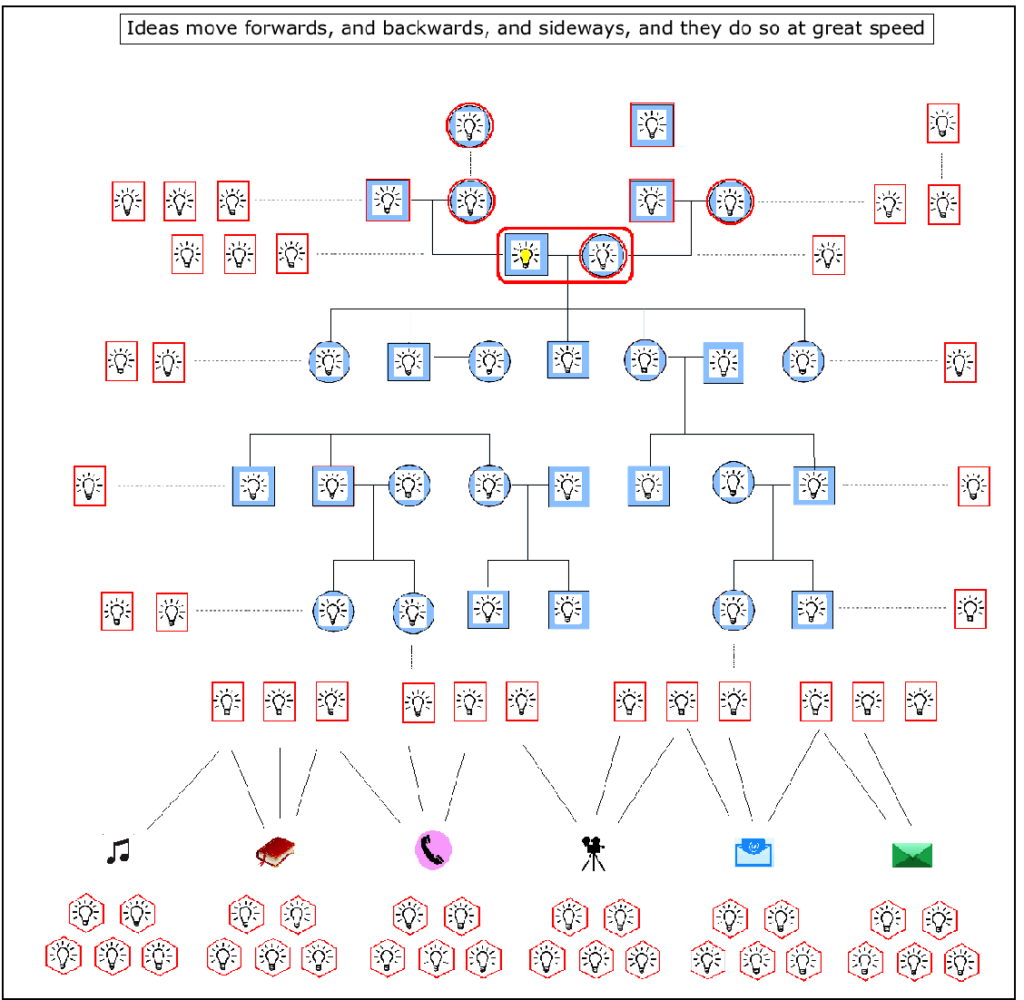
In fact, given the colourful and substantial nature of human language, particularly with all the practical and technological follow ons just mentioned, it is easy to understate, or even forget the primary role of the imagination in the generation of new ideas. Yet it is in the creation of new ideas that we see the emergence and proliferation of Social Space. New ideas, new designs, and for that matter, even new languages all come from our ability to imagine how things can and might be – the imagination always comes first. And although language frees the solitary imagination from its own individual and limited lifetime, it is the imagination and only the imagination that creates all the meaning that makes up the entirety of our lives. Which means we have to give it primary billing in our assessment of what turned ape to human, closely shadowed by the language that subsequently empowers it to create what we now call our culture. Or to put it another and more picturesque way, it was the evolution of the imagination, feathered by language, that gave us the wings on our heads.
Nevertheless, we are bound to ask how such a risky thing as the imagination evolved in the first place. How could a system that makes things up, seemingly ‘as it goes along’ succeed in evolving from a system that maintains such an accurate and literal fix on natural reality as does animal nature? Well, one answer is that although the chimp departs from the literal world of ‘what is’ when it imagines different ways of getting the banana, this freedom nevertheless begins and ends in the needs of the animal. The chimp is taking a minimal risk because both its recognition and response to the banana are held firmly in place by the directives of Genetic Space. (There’s a banana. Go get it now!).
The problem about the imagination is that the idea of ‘what might be the case’ represents an altogether different order of commitment to physical reality than the ‘what actually is, or has been the case’ of the animal kingdom. Different because speculation about what may or may not be the case can take untried and untested options seriously to such an extent that it looks more like a form of gambling than a serious behavioural strategy created by millions of years of evolution. So we have to ask ourselves how such a strange commitment as this could survive at all, especially in the company of all of those tried and tested evolutionary strategies that have been honed to such a pitch of perfection over so many millennia?
Perhaps a good way of looking at this might be to compare the use of the imagination with the use of mind bending drugs. The kind of drug induced vagaries of thought that can afflict us when we for example ‘drop acid’ can have very strange effects on our perception. And once the mind is doing its own thing, and ignoring the reality outside its organic housing, its owner is in real danger of damage or death. Standing on a parapet, in the daze induced by taking ketamines, and jumping off because you suddenly ‘know you can fly’ is an example of how the imagination can go badly wrong if not controlled properly. Indeed, it may be that the imagination only has to take us slightly out of tune with reality for numerous risks to emerge. After all, a monkey, about to leap to another branch in the high canopy of the forest has to get it exactly right, or it will plunge to the ground far below. And if it has any false promptings, based on putting the imagined picture before the real time picture, then these could be fatal. Which must mean that the environment has zero tolerance for the whimsy of altered perceptions of the kind likely to emerge from the use of an unfettered imagination. All of which suggests that the evolution of the imagination must be accompanied by a parallel ‘discipline of mind’ that keeps its user from veering off the paths of phylogenetic and ontogenetic reality, and into the jaws of death.
Given that evolution has hardwired a number of basic drives within our body, including various fears and hungers that are of great importance to us in our orientation to the world outside, it seems likely that these drives have kept pace with the evolution of the imagination, and have channelled its potential follies into a survival bound direction. Certainly these drives must be keeping the human imagination within quite tight boundaries, or we would presumably go ‘off the rails’ and die before passing on the genes that support such potentially dangerous flights of fantasy. So the story of just how the risks incurred by imaginative thought have been avoided in our evolution is going to be every bit as fascinating to read as the one about how ‘picture matching’ evolved. But we have to leave them both at this point. Leave them because we must stay loyal to the intent of this section, which is merely to propose the question of where the imagination might belong in the evolution of animal behaviour.
In summary then, in the three parts of this section, we have gone ‘Off the Map’ of the hard sciences, and their already well charted physical and living landscapes, into the uncharted territory of human meaning. With Bugs Bunny, we have gone off the cliff, and defied gravity, to reveal a little about third level logic. We then returned to earth to explain more fully the place of Social Space in the scheme of things, going off planet to show that the three levels of reality are rather more than just an arbitrary distinction. Then we speculated about our ability to do just precisely that, and discussed how the imagination fits into the rest of animal grammar. The role of the twin agency of the hand and the mouth in relation to the evolution of the imaginations was then looked at. And once these two had released the imagination from its otherwise smothering isolation and physical impotency, it was free to turn the ‘might’ of the subjunctive tense into the very real ‘Might’ of the appliance of crafts and then science. At which point, Social Space bloomed, and the human race became the significant level of reality that we know it to be today.
This brings us to the point where we move away from the insights of biology, and into the relatively unexplored territory of Social Space. The question is this: In our own lives, how much of what we do and think is controlled by animal logic, and how much is the result of a very different and indeed social logic? Or to put it another way, can we explain human thought and action through the same biological insight that we use to explain chimp behaviour? Well, the logic of the joke is a prime example of a unit of human meaning where it is very hard to see how the theories of biology can possibly shed any real light on what we find funny. Because although the physical reaction of laughter or smiling has a basis in our primate origins, the semantic complexity of even the simplest joke requires, as we shall see, a brand new set of understandings that owes very little to those that illuminate the nature of Genetic Space. And a new set of understandings that must lead to a new science that goes far beyond biology. Far beyond biology that is, to a science of meaning.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}